InVivoMAb anti-human CD28
Product Details
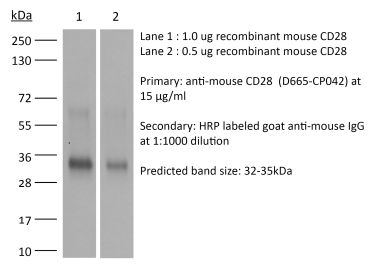
The 9.3 monoclonal antibody reacts with human CD28, a 45 kDa costimulatory receptor and a member of the Ig superfamily. CD28 is expressed by thymocytes, most peripheral T cells, and NK cells. CD28 is a receptor for CD80 (B7-1) and CD86 (B7-2). Signaling through CD28 augments IL-2 and IL-2 receptor expression as well as cytotoxicity of CD3-activated T cells. The 9.3 antibody has been shown to stimulate the proliferation of human T cells in vitro.Specifications
| Isotype | Mouse IgG2a |
|---|---|
| Recommended Isotype Control(s) | InVivoMAb mouse IgG2a isotype control, unknown specificity |
| Recommended Dilution Buffer | InVivoPure pH 7.0 Dilution Buffer |
| Conjugation | This product is unconjugated. Conjugation is available via our Antibody Conjugation Services. |
| Immunogen | Human T lymphocytes |
| Reported Applications | in vitro T cell stimulation/activation |
| Formulation |
PBS, pH 7.0 Contains no stabilizers or preservatives |
| Endotoxin |
<2EU/mg (<0.002EU/μg) Determined by LAL gel clotting assay |
| Purity |
>95% Determined by SDS-PAGE |
| Sterility | 0.2 µm filtration |
| Production | Purified from cell culture supernatant in an animal-free facility |
| Purification | Protein G |
| RRID | AB_2687729 |
| Molecular Weight | 150 kDa |
| Storage | The antibody solution should be stored at the stock concentration at 4°C. Do not freeze. |
Recommended Products
-

Recommended Isotype Control(s)
InVivoMAb mouse IgG2a isotype control, unknown specificity
-

Recommended Dilution Buffer
InVivoPure pH 7.0 Dilution Buffer
in vitro T cell stimulation/activation
Huang, X., et al. (2021). "DNA scaffolds enable efficient and tunable functionalization of biomaterials for immune cell modulation" Nat Nanotechnol 16(2): 214-223. PubMed
Biomaterials can improve the safety and presentation of therapeutic agents for effective immunotherapy, and a high level of control over surface functionalization is essential for immune cell modulation. Here, we developed biocompatible immune cell-engaging particles (ICEp) that use synthetic short DNA as scaffolds for efficient and tunable protein loading. To improve the safety of chimeric antigen receptor (CAR) T cell therapies, micrometre-sized ICEp were injected intratumorally to present a priming signal for systemically administered AND-gate CAR-T cells. Locally retained ICEp presenting a high density of priming antigens activated CAR T cells, driving local tumour clearance while sparing uninjected tumours in immunodeficient mice. The ratiometric control of costimulatory ligands (anti-CD3 and anti-CD28 antibodies) and the surface presentation of a cytokine (IL-2) on ICEp were shown to substantially impact human primary T cell activation phenotypes. This modular and versatile biomaterial functionalization platform can provide new opportunities for immunotherapies.
in vitro T cell stimulation/activation
Koh, J., et al. (2020). "MDSC subtypes and CD39 expression on CD8(+) T cells predict the efficacy of anti-PD-1 immunotherapy in patients with advanced NSCLC" Eur J Immunol 50(11): 1810-1819. PubMed
The major suppressive immune cells in tumor sites are myeloid derived suppressor cells (MDSCs), tumor-associated macrophages (TAMs), and Treg cells, and the major roles of these suppressive immune cells include hindering T-cell activities and supporting tumor progression and survival. In this study, we analyzed the pattern of circulating MDSC subtypes in patients with non-small cell lung cancer (NSCLC) whether those suppressive immune cells hinder T-cell activities leading to poor clinical outcomes. First, we verified PMN-MDSCs, monocytic-MDSCs (M-MDSCs), and Treg cells increased according to the stages of NSCLC, and MDSCs effectively suppressed T-cell activities and induced T-cell exhaustion. The analysis of NSCLC patients treated with anti-PD-1 immunotherapy demonstrated that low PMN-MDSCs, M-MDSCs, and CD39(+) CD8(+) T cells as an individual and all together were associated with longer progression free survival and overall survival, suggesting PMN-MDSCs, M-MDSCs, and CD39(+) CD8(+) T cells frequencies in peripheral blood might be useful as potential predictive and prognostic biomarkers.
in vitro T cell stimulation/activation
Hill, E. V., et al. (2015). "Glycogen synthase kinase-3 controls IL-10 expression in CD4(+) effector T-cell subsets through epigenetic modification of the IL-10 promoter" Eur J Immunol 45(4): 1103-1115. PubMed
The serine/threonine kinase glycogen synthase kinase-3 (GSK3) plays an important role in balancing pro- and anti-inflammatory cytokines. We have examined the role of GSK3 in production of IL-10 by subsets of CD4(+) T helper cells. Treatment of naive murine CD4(+) T cells with GSK3 inhibitors did not affect their production of IL-10. However, treatment of Th1 and Th2 cells with GSK3 inhibitors dramatically increased production of IL-10. GSK3 inhibition also led to upregulation of IL-10 among Th1, Th2, and Th17 subsets isolated from human blood. The encephalitogenic potential of GSK3 inhibitor treated murine Th1 cells was significantly reduced in adoptive transfer experiments by an IL-10-dependent mechanism. Analysis of the murine IL-10 promoter in response to inhibition of GSK3 in Th1 cells showed modification to a transcriptionally active state indicated by changes in histone H3 acetylation and methylation. Additionally, GSK3 inhibition increased expression of the transcription factors c-Maf, Nfil3, and GATA3, correlating with the increase in IL-10. These findings are important in the context of autoimmune disease since they show that it is possible to reprogram disease-causing cells through GSK3 inhibition.
in vitro T cell stimulation/activation
Bushkin, Y., et al. (2015). "Profiling T cell activation using single-molecule fluorescence in situ hybridization and flow cytometry" J Immunol 194(2): 836-841. PubMed
Flow cytometric characterization of Ag-specific T cells typically relies on detection of protein analytes. Shifting the analysis to detection of RNA would provide several significant advantages, which we illustrate by developing a new host immunity-based platform for detection of infections. Cytokine mRNAs synthesized in response to ex vivo stimulation with pathogen-specific Ags are detected in T cells with single-molecule fluorescence in situ hybridization followed by flow cytometry. Background from pre-existing in vivo analytes is lower for RNAs than for proteins, allowing greater sensitivity for detection of low-frequency cells. Moreover, mRNA analysis reveals kinetic differences in cytokine expression that are not apparent at the protein level but provide novel insights into gene expression programs expected to define different T cell subsets. The utility of probing immunological memory of infections is demonstrated by detecting T cells that recognize mycobacterial and viral Ags in donors exposed to the respective pathogens.
in vitro T cell stimulation/activation
Kovacs, B., et al. (2005). "Ligation of CD28 by its natural ligand CD86 in the absence of TCR stimulation induces lipid raft polarization in human CD4 T cells" J Immunol 175(12): 7848-7854. PubMed
Stimulation of resting CD4 T cells with anti-CD3/CD28-coated beads leads to rapid polarization of lipid rafts (LRs). It has been postulated that a major role of costimulation is to facilitate LR aggregation. CD86 is up-regulated or expressed aberrantly on immune cells in a wide array of autoimmune and infectious diseases. Using an Ig fusion with the extracellular domain of CD86 (CD86Ig) bound to a magnetic bead or K562 cells expressing CD86, we demonstrated that ligation of CD28 by its natural ligand, but not by Ab, induced polarization of LRs at the cell-bead interface of fresh human CD4 T cells in the absence of TCR ligation. This correlated with activation of Vav-1, increase of the intracellular calcium concentration, and nuclear translocation of NF-kappaB p65, but did not result in T cell proliferation or cytokine production. These studies show, for the first time, that LR polarization can occur in the absence of TCR triggering, driven solely by the CD28/CD86 interaction. This result has implications for mechanisms of T cell activation. Abnormalities in this process may alter T and B cell tolerance and susceptibility to infection.